Wize University Biochemistry Textbook > Eukaryotic Transcription
Eukaryotic Transcription

0:00 / 0:00
Transcription in Eukaryotes
Transcription in eukaryotic cells is fundamentally the same as that process in prokaryotic cells. Here we will point out some more key differences between the two.
- Eukaryotic DNA is packaged into chromatin, which in the condensed state (default state) results in a promoter that is inaccessible to RNA polymerase. Transcription can only occur when DNA is decondensed and dissociated from the histones around which it is wound.
- Transcription and translation cannot occur at the same time because eukaryotes have a separate compartment for the DNA (nucleus).
- Prokaryotic cells: genes with common function are arranged linearly and are transcribed together on a single mRNA (polycistronic).
- Eukaryotic cells: genes with common functions can be scattered throughout the genome, on different chromosomes and can therefore be transcribed and regulated separately from one another.
- Eukaryotic mRNA is always monocistronic: one mRNA = one protein.
- Eukaryotes have three types of RNA polymerase:
- RNAP I: transcribes rRNA
- RNAP II: transcribes mRNA
- RNAP III: transcribes tRNA
- Messenger RNA requires extensive modification before being translated into protein (maturing of RNA).
- Promoters are more complex but also have a consensus sequence similar to that in prokaryotes. This is called the TATA box and is located about -30 nucleotides upstream of the transcriptional start site.
- RNA polymerase needs the help of transcription factors to bind to DNA and start transcribing. Instead of a simple sigma factor, several transcription factors (TFs) bind to the promoter to recruit RNA Polymerase II. See below for more details on this.

8. Elongation is similar to prokaryotes but termination can be different (depends on RNA Polymerase type).
Transcriptional Control
- The equivalent of sigma proteins in prokaryotic transcription are basal transcription factors in eukaryotic cells.
- Basal transcription factors include a TATA-binding protein (TBP) that binds to the TATA box in the promoter of eukaryotic genes. Following TBP binding, the RNA polymerase is recruited.
- Other transcription factors (such as enhancers) bind to different areas within the gene and interact with the polymerase to initiate transcription.
- Enhancers can even be located far away from the genes they affect.
- Transcription factors help to coordinate gene expression. Genes located on different chromosomes can be controlled simultaneously through the use of the same transcription factor.


0:00 / 0:00
General Transcription Factors Required for Initiation
Phosphorylation of the RNAP II carboxy-terminal domain (CTD)
- The CTD is made up of Tyr-Ser-Pro repeats (52 repeats in humans)
- During initiation of transcription, the CTD is unphosphorylated
- As the polymerase begins the elongation phase of transcription, the CTD is phosphorylated
- RNA processing proteins, like capping, elongation, and splicing factors can bind to the CTD depending on it's phosphorylation state
General Transcription Factors (GTFs)
Transcription Factor II (TFII) is made up of multiple subunits which help position RNAP II at the start site and initiate transcription
- TFIID consists of a TATA box Binding Protein (TBP) and 13 other TBP-Associated Factors (TAFs)
- TFIID forms a complex with TFII A and TFIIB and the TATA-box
- TFIIF and RNAP II then bind
- TFIIE binds and creates a site for TFIIH to bind
- TFIIH has helicase activity which unwinds the DNA
- After transcription has been initiated, TFIIH has kinase activity which phosphorylates the CTD of RNAPII

0:00 / 0:00
Regulation of Eukaryotic Transcription
TRANSCRIPTIONAL ACTIVATORS
- Activate transcription by binding to control regions on the DNA (enhancer sites, promoters, initiator sites, etc.)
- Transcriptional activators have a DNA binding domain and an activation domain
- DNA binding domain: Zinc finger domains (C2H2 or C4), leucine zipper, basic Helix-Loop-Helix
- Activation domains: short structures containing high percentage of one or two amino acids (e.g. Asp, Glu, Gln, Pro, Ser or Thr). Facilitates protein-protein interactions: may have a random structure until interaction with a co-activator protein
Exam Tip
General Transcription Factors (GTFs) are a unique set of transcription factors because they are required to initiate almost all RNAP II activity at the promoter. GTFs are NOT considered to be activators of transcription in the same way as the transcription factors discussed in this lesson.
TRANSCRIPTIONAL REPRESSORS
- Repress transcription by binding to control regions on the DNA
- Transcription repressors have a DNA binding domain and a repression domain
- DNA binding domain: Zinc finger domains (C2H2 or C4), leucine zipper, basic Helix-Loop-Helix
- Repression domain: short structures (<15 amino acids). Facilitates protein-protein interactions: may need to bind to co-repressor proteins to silence transcription
How does a finite set of transcription factors control the expression of so many genes??
Wize Tip
Control regions on DNA often have multiple sites where transcriptional activators AND repressors can bind. Depending on which type of transcription factor binds to the control region, gene transcription can be activated or repressed.
- Heterodimers: combinations of different transcriptions binding together to interact with different DNA sequences.
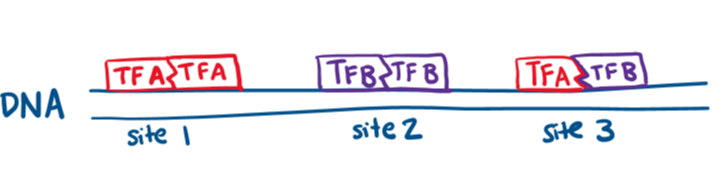
- Inhibitory factors: Can block the activity of some transcription factor DNA binding domains
- Cooperative Binding: Unrelated transcription factors binding to neighboring sites
- Example: A transcription factor with weak repression activity binds nearby another weak repressor= STRONG repression
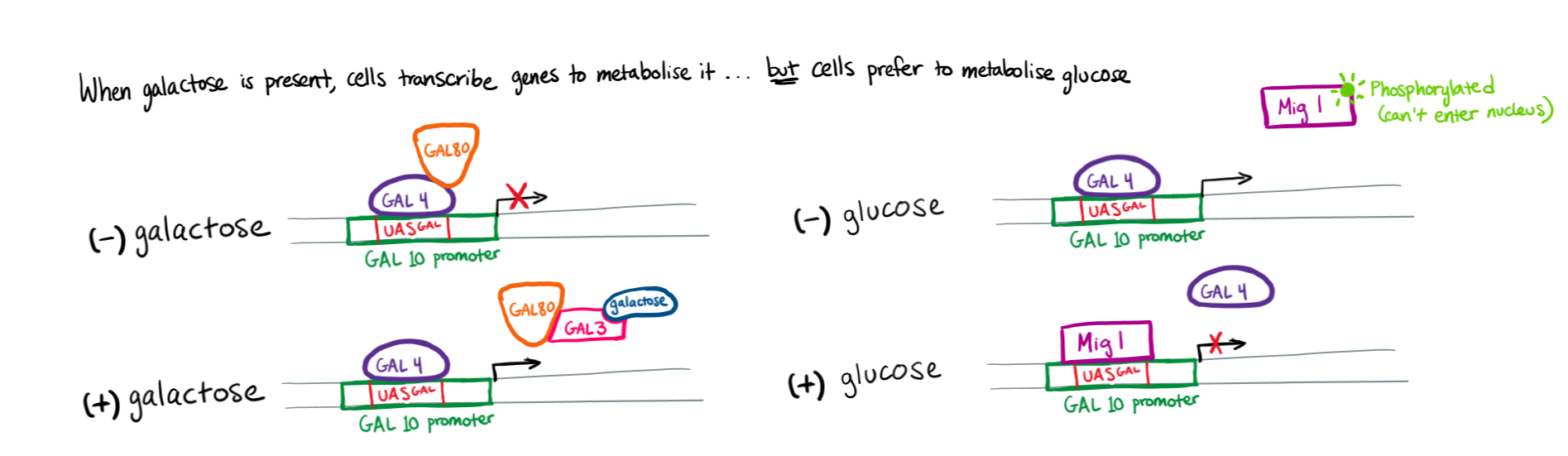
Example of Eukaryotic transcriptional Control: GALACTOSE METABOLISM

0:00 / 0:00
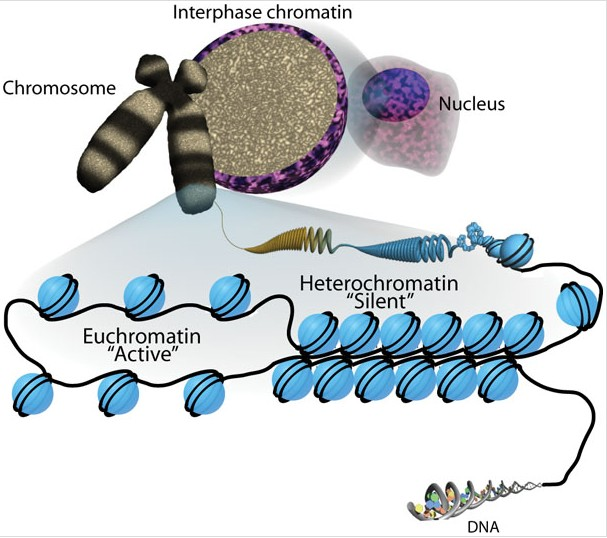
Heterochromatin vs. Euchromatin
DNA replication and transcription is dependent on what physical state chromatin is in.
HETEROCHROMATIN is the tightly packed, condensed form of chromatin- transcriptionally "silent"
EUCHROMATIN is more open, de-condensed form of chromatin- transcriptionally "active"
Post-translational Histone Modification
The C-terminus and N-terminus tails of the histone proteins can be modified
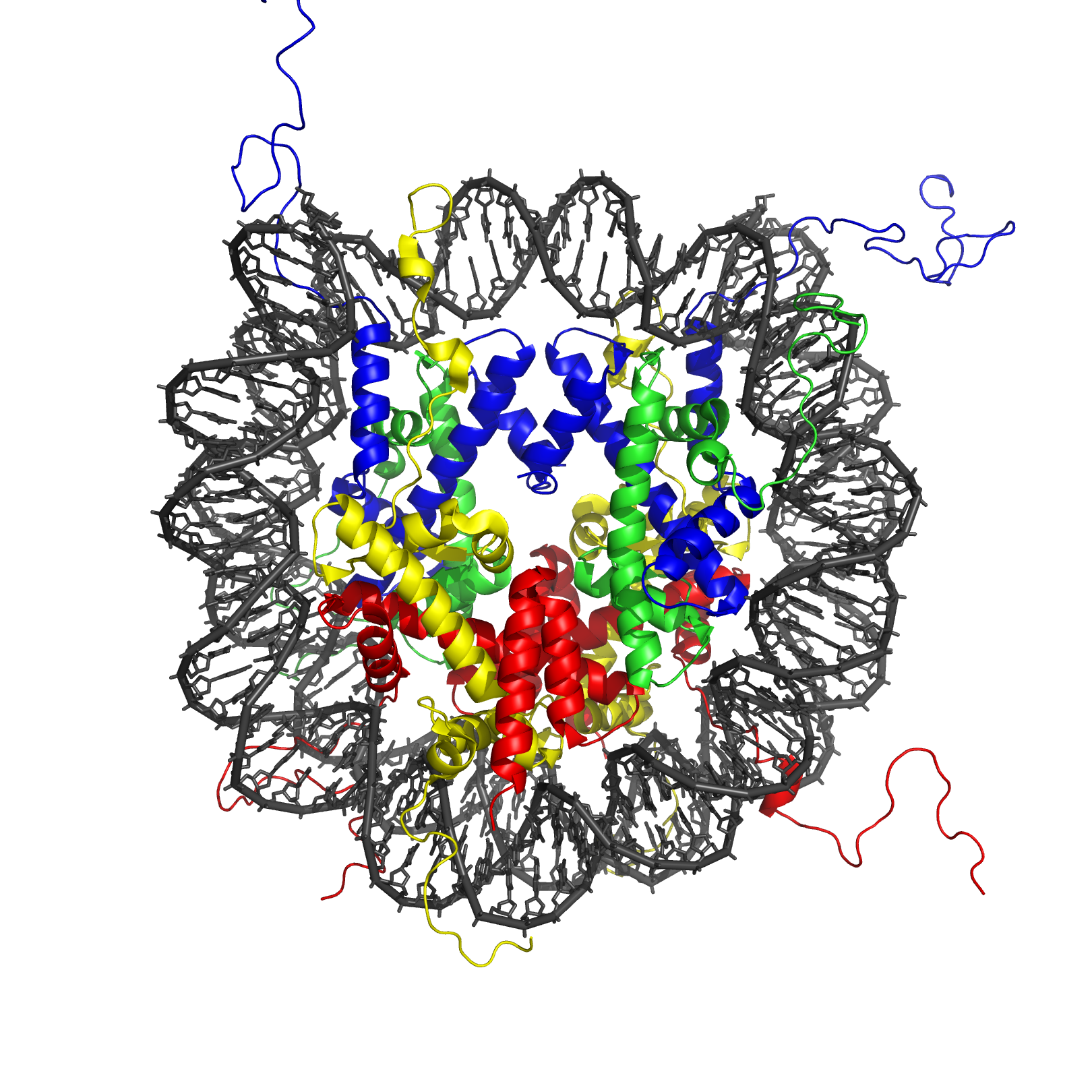
The most widely-studied post-translational modifications are:
- mono-, di- or tri- methylation on Lysine (K) or Arginine (R)
- acetylation on lysine (K)
- Phosphorylation on Serine (S) or Threonine (T)
Some of these modifications correlate to chromatin state. For example:
- H3K4me3 causes chromatin to decondense= euchromatin
- H3K9me2 causes chromatin to condense= heterochromatin
- Acetylation of lysine usually correlates with euchromatin where as deacetylation correlates with heterochromatin
POSITIVE FEEDBACK LOOP: Once heterochromatin begins, chromatin condensation can spread. A methylation mark can recruit proteins (e.g. HP1) to the DNA which, in turn, recruit methyl transferases.
Similarly, once euchromatin begins, chromatin de-condensation can spread. An acetylation mark can recruit proteins (e.g. bromodomain proteins) to the DNA, which, in turn, recruits acetylases.